

Сложно себе представить новогодний интерьер или даже экстерьер без главного атрибута зимнего праздника — новогодней елки. Будучи вечнозелеными, хвойные деревья стали символом жизни, что не удивительно, ведь подавляющее большинство других растений в зимний период либо погибают, либо сбрасывают листву. Ученым давно известна разница между лиственными и хвойными деревьями, но некоторые моменты доселе оставались загадкой. Ученые из университета Умео (Швеция) провели исследование, в котором установили причину «вечнозелености» хвойных деревьев. Какие процессы поддерживают внешний вид хвои вне зависимости от сезона, насколько они сложны, и как это повлияло на людей? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.

Основа исследования
Фотосинтез — термин, знакомый нам со школьной скамьи. Данный процесс является основой многих форм жизни на Земле, позволяя им расти за счет солнечной энергии. Эта удивительная способность позволила многим растениям в ходе эволюции выработать множество различных методик ее реализации в зависимости от среды произрастания.
Одной из таких сред являются бореальные леса, занимающие примерно 70% от всех хвойных лесов планеты. Данная экосистема территориально связана с регионами, где преобладает холодный, умеренно холодный и умеренный климат. В таких лесах присутствует довольно мало видов растений, а большую часть составляют вечнозеленые хвойные породы, такие как сосна и ель.
Особенностью холодных регионов с точки зрения растений является короткий вегетационные периоды и холодные, снежные зимы. Следовательно, факт наличия в столь малоприятных условиях хвойных растений должен означать наличие у них некого механизма приспособления, основанного на регуляторных процессах, которые защищают фотосинтетический аппарат в их иглах от воздействия окружающей среды.

Однако четкого понимания того, что лежит в основе этого механизма нет. Существует несколько теорий, основанных на многочисленных исследованиях. В ходе одного из них было установлено, что хвойные иголки зимой переходят в состояние длительного тушения*.
Нефотохимическое тушение* (или NPQ от non-photochemical quenching) — механизм защиты фотосинтетического аппарата от света высокой интенсивности. Процесс тушения заключается в поглощении избыточной энергии синглетного возбужденного хлорофилла молекулой-акцептором с последующим переходом этой молекулы в основное энергетическое состояние при помощи усиленной внутренней конверсии.Также было установлено наличие изменений в организации и составе антенн фотосистемы II (PSII от photosystem II). При этом ученые не отбрасывают и гипотетическую важность фотосистемы I (PSI от photosystem I) в процессе нефотохимического тушения поглощенного света.
Параллельно, по мнению ученых, могут действовать несколько защитных механизмов, и они, вероятно, активируются задолго до возникновения экстремального стресса (т. е. до холодов). Кроме того неясна ситуация весной, когда все еще холодно, поэтому биохимическая и метаболическая активность растений сильно ограничена, но солнечная радиация уже достаточно высока. Следовательно, механизм защиты должен быть активирован до зимы, потом поддерживаться на нужном уровне в течение зимних холодов, а потом отключаться, чтобы фотосинтетический аппарат восстановился и летом достиг состояния активного роста.
В ранее проведенных исследованиях у некоторых зимующих хвойных растений было замечено значительное падение фотохимии фотосистемы II (измеряемой как Fv/Fm). На основе этого наблюдения было выдвинуто объяснение: разупаковка тилакоидных* мембран и связанные с этим изменения ультраструктуры хлоропластов в соответствии с падением максимальной флуоресценции PSII.
Тилакоид* — ограниченный мембраной компартмент внутри хлоропластов и цианобактерий, в котором протекают светозависимые реакции фотосинтеза.У высших растений стэкинг и гетерогенность тилакоидов играют решающую роль в локализации PSII в гранах (плотно прижатые слои тилакоидов) и PSI в ламеллах стромы.
В рассматриваемом нами сегодня труде ученые попытались отследить стационарную флуоресценцию хлорофилла, сверхбыструю флуоресценцию с временным разрешением и ультраструктуру хлоропластов в хвое сосны обыкновенной с осени до лета в течение трех лет.
Результаты исследования показали, что тушение флуоресценции хлорофилла и разрушение тилакоидов тесно связаны, взаимозависимы и имеют решающее значение для выживания вечнозеленых хвойных деревьев в условиях экстремальной бореальной зимы и ранней весны, когда температуры низкие, но уровни солнечной радиации могут быть высокими.
Результаты исследования
Чтобы отслеживать эффективность фотосинтеза, были записаны некоторые параметры PSII и PSI в течение трех последовательных сезонов роста (2015–2016, 2016–2017 и 2017–2018), а также сопутствующие изменения суточной температуры воздуха и солнечной радиации (1a-1c).

Изображение №1
Как поясняют авторы исследования, для простоты на основных изображениях представлены данные только с 2017 по 2018 год, которые были разделены на пять сезонов в зависимости от погодных параметров: лето (S, июнь – август), осень (A, сентябрь – середина ноября), зима (W, середина ноября – середина февраля), ранняя весна (ES, середина февраля – середина апреля) и поздняя весна (LS, середина апреля – июня).
Были обнаружены характерные сезонные закономерности максимальной флуоресценции PSII (Fm) и максимальной квантовой эффективности PSII (выраженной как Fv/Fm). Fv/Fm было самым высоким летом, затем падало с понижением температуры окружающей среды во время осени и зимы (1d и 1e) и было самым низким ранней весной (на 63% ниже, чем летом), когда низкие температуры совпадали с повышением солнечной радиации (1a-1c). В период поздней весны Fv/Fm постепенно увеличивался с повышением температуры и достигал максимума в летний период. Важно и то, что на сезонные изменения, зарегистрированные в Fv/Fm, больше всего влияют изменения Fm (1d), а не Fo (основная собственная флуоресценция).
Для более глубокого понимания работы PSII была измерена доля поглощенной световой энергии (1f), используемой фотохимией PSII (ΦPSII). Так зимой ΦPSII значительно снизился и достиг минимальных значений (19% от значений летом) в период ранней весны.
Далее была проведена количественная оценка поглощенной световой энергии, термически рассеиваемой нефотохимическим тушением (NPQ).
Компонент NPQ, который играет решающую роль в условиях переменного освещения, это быстрый компонент (ΔpH-PsbS-зависимый-qE3 или зеаксантин-зависимый-qZ4), который увеличивается с ростом интенсивности света во все сезоны. Летом и осенью быстрый компонент не достигал стационарной фазы даже при освещении 1500 мкмоль/м2/с (т.е. мкмоль фотонов на квадратный метр в секунду — фотосинтетически активная радиация). Но вот зимой и поздней весной стационарная фаза достигалась при 500, а ранней весной при 300 мкмоль/м2/с (1g). Важнее то, что ранней весной NPQ был намного медленнее (на ~ 50% меньше, чем летом) из-за меньшей амплитуды быстрого компонента (1h). Это объясняется тем, что ранней весной иглы уже имеют статический NPQ.
Но вот квантовый выход нерегулируемых и/или конститутивных потерь (ΦNO) энергии был высоким во время именно ранней весной (1i). Это говорит о том, что данный статический NPQ представляет собой часть поглощенной световой энергии, которая не влияет на фотохимию (ΦPSII) и не рассеивается термически в результате быстро регулируемых процессов NPQ (qE/qZ).
Для подтверждения того, что статическое тушение является проявлением длительного тушения, иглы хвои ранней весны искусственно расслаблялись (восстановительный процесс) в течение 48 часов при слабом освещении (80 мкмоль/м2/с) с фотопериодом 18/6 часов (образцы ER — хвоя после восстановления). Во время этого процесса наблюдалось следующее: в первые 6-8 часов восстановление было незначительным (менее 10%), спустя 24 часа — 30% восстановление (1j), а спустя 48 часов иглы восстановились почти полностью (95%) до уровня игл весеннего периода.
Fo существенно не изменилось в течение периода восстановления (1j), но изменения в Fm следовали той же динамике восстановления, что и Fv/Fm, хотя максимальная флуоресценция (Fm) была намного выше у весенних игл, чем опытных образцах спустя 48 часов искусственного восстановления.
ΦPSII не показывал значительных изменений в течение первых 12 часов восстановления, однако спустя 24 часа показал 50% от уровней весенних игл и 90% спустя 48 часов (1k). qE/qZ-зависимая кинетика быстрого компонента NPQ показывала аналогичную ΦPSII схему восстановления (1l).
Важно и то, что светозависимая индукция быстрого компонента NPQ (qE/qZ) была медленнее в образцах ER (опытные образцы), чем в образцах S (взятые в природе летом), даже после 48 часов восстановления. Это, скорее всего, связано с фотоингибированием.

Изображение №2
Сезонные изменения в распределении поглощенной световой энергии внутри PSI показали, что Y(I), т.е. квантовый выход, был самым высоким летом, уменьшался осенью и зимой, а минимальных значений достигал ранней весной (2а). Несмотря на то, что Y(I) уменьшился, он подвергался гораздо меньшему влиянию, чем активность PSII (ΦPSII) во время холодных периодов (1f). Y(NA), т.е. рассеяние энергии из-за ограничения на акцепторной стороне, следовало той же сезонной модели с минимальными значениями, зарегистрированными ранней весной, и длительным восстановлением до лета (2b).
В отличие от Y(NA), Y(ND), т.е. рассеяние энергии из-за ограничения на донорной стороне, значительно увеличивалось в холодные периоды и достигало максимума (на 69% выше, чем весной) ранней весной, а затем постепенно снижалось до минимума летом (2с).
Данные наблюдения могут говорить о том, что ранней весной большая часть поглощенной световой энергии рассеивается окисленным донором P700 (P700+), чтобы предотвратить фотоингибирование, вызванное чрезмерным восстановлением акцепторной стороны при повреждении кластеров железо/сера.
В течение первых 3 часов периода восстановления ΦPSI существенно не изменился, но постепенно вернулся к летним уровням в течение 48 часов (2d). Y(NA) не показал значительных изменений во время восстановления (2е). А вот Y(ND) несколько увеличивался в течение первых 3 часов, но затем постепенно снижался.
На основании этих результатов можно сделать вывод, что восстановление эффективности использования света PSI занимает более 48 часов (2f).
Далее, чтобы выяснить механизм «длительного тушения», наблюдаемого у сосны в ранние весенние периоды, были выполнены сверхбыстрые измерения флуоресценции с временным разрешением на неповрежденной хвое сосны. Дополнительно использовалась специально разработанная камера хранения образцов, где поддерживалась температура -20 °C.
Анализ флуоресценции образцов показал ожидаемые результаты: летние иглы вообще не содержали «длительного тушения», тогда как весенние показали самые высокие уровни «длительного тушения».
Чтобы выяснить механизмы этого тушения, следы затухания флуоресценции летних игл в исходном состоянии (3а) сравнивали с иглами, собранными ранней весной, и иглами после искусственного восстановления при комнатной температуре (ER).
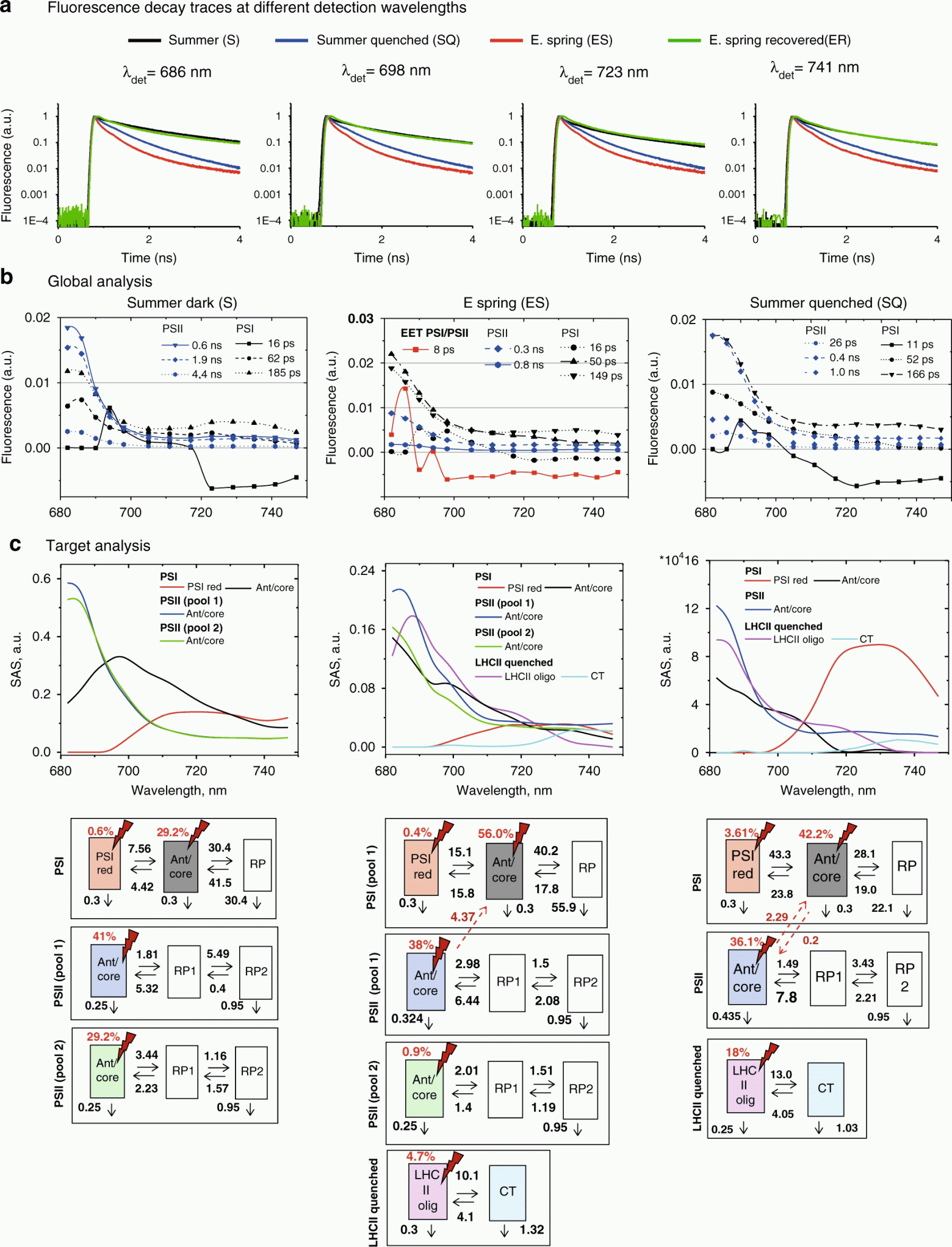
Изображение №3
Чтобы понять, происходит ли длительное тушение, наблюдаемое ранней весной, посредством механизма, аналогичного светоиндуцированному тушению, также были измерены летние иглы, подвергнутые воздействию сильного освещения в течение 30 минут (SQ — летнее тушение). Как показывает прямое сравнение кривых затухания (3а), флуоресценция в SQ иглах намного короче, чем у летних (S) игл.
Однако затухание флуоресценции игл ранней весны (ES) по-прежнему значительно короче, чем у образцов SQ, что характеризует «длительное тушение» в ES образцах как наиболее выраженное тушение при всех длинах волн и во всех условиях.
Напротив, образцы ER и S (летние) показали очень похожее затухание флуоресценции. Это указывает на то, что образец ES достаточно (хотя и не полностью) восстановился после длительного тушения в течение 48 часов после восстановительной обработки.
Чтобы получить первые подсказки о механизме, лежащем в основе длительного тушения в начале весны, был проведен глобальный анализ всех наборов данных: иглы ES, S, SQ (3b) и ER.
Для соответствия кинетике флуоресценции во всех четырех случаях требовалось шесть спектров, связанных с распадом (DAS). Три DAS были предварительно отнесены к PSI: 11–16 пс, 50–60 пс, 150–180 пс (3b). Самый быстрый компонент представляет собой энергетическое равновесие между Chlred (хлорофилл красный) и Chlblue (хлорофилл синий). Второй DAS в летних иглах имеет максимум около 685–690 нм с широкой эмиссией в области 700–720 нм. Следовательно, он в основном представляет распад ядра PSI в сочетании с 710–720 хлорофилл антенных комплексов. Третий DAS (185 пс) имеет максимум при 680 нм с некоторым вкладом при 730 нм, и поэтому его следует в первую очередь отнести к LHCII (антенный комплекс II) и некоторому количеству Chlred в PSI.
В состояниях S и ER оставшиеся три пика DAS находятся при 682 нм и, следовательно, относятся к PSII. Их время жизни составляет 0.5–0.6 нс, 1.5–2.0 нс и 4 нс, аналогично тому, что было выявлено ранее для PSII в закрытом состоянии. Однако в ES наблюдались два значительно более короткоживущих DAS, связанных с PSII, с временами жизни 0.3 и 0.8 нс. Это демонстрирует, что кинетика PSII действительно подвергается тушению в ES иглах.
Колебания в спектрах обусловлены смешиванием составляющих компонентов, представляющих перенос энергии возбуждения (EET) от пулов хлорофилла типа a, излучающих при ~680 нм, к пулам, излучающим при 690 нм и 700-730 нм. Это говорит о том, что в иглах ранней весны происходит эффективный перенос энергии возбуждения между PSII и/или LHCII и PSI.
Другими словами, результаты глобального анализа предполагают, что длительное тушение в ES состоянии (ранней весной) включает как PSII/LHCII, так и PSI посредством прямой передачи энергии. Этот механизм обеспечивает гораздо более сильное тушение, чем тушение в летний период, вызванное сильным светом.
Для лучшего понимания происходящих процессов и механизмов, с ними связанных, ученые также провели моделирование кинетики флуоресценции хвои, которая не испытывала тушение. В данном целевом анализе конкретная кинетическая модель проверяется с использованием линейной, неизменной во времени, компартментной модели. На изображении 3с каждый из компартментов представлен квадратной рамкой. Каждая рамка относится к любому из трех различных компонентов: PSI, PSII и LHCII после тушения. Компартменты, излучающие флуоресценцию, представлены цветными прямоугольниками. Красные молнии и цифры рядом с ними представляют так называемый вектор возбуждения, указывающий процент возбуждений, направленных на каждый компартмент, и отражающий относительное количество Chl a (то есть относительный размер Chl a антенны) в нем. Бесцветные рамки представляют собой неизлучающие компартменты, такие как радикальные пары (RP), которые вносят вклад в общую наблюдаемую кинетику.
В летних иглах кинетика PSI описывалась двумя излучающими компартментами (PSI красный, Ant/core) и одной парой неизлучающих радикалов (RP). Кинетика комплекса антенна PSII/RC описывалась одним излучающим компартментом (Ant / core) и двумя неизлучающими радикальными парами (RP1 и RP2).
Для достижения необходимого соответствия в данном анализе необходимо было два пула комплексов PSII с разными скоростями распада (пул 1 и пул 2) для летнего образца. Эти два пула различаются константами скорости разделения зарядов, что типично для PSII частиц с разными размерами антенн, что приводит к разному среднему времени жизни.
В летнем образце хвои на PSI приходится 30% полного сечения поглощения на длине волны возбуждения (662 нм), тогда как на два пула PSII приходится 40% и 30% от общего сечения поглощения. Никакого внутреннего тушения, связанного с каким-либо процессом NPQ (нефотохимическое тушение) в этих PSII компартментах, не наблюдалось.
Модель для образцов ранней весны весьма схожа с моделью летней хвои. Основное отличие состоит в том, что меньший PSII пул (с очень маленьким поперечным сечением поглощения, около 10%) показывает необычно малую скорость разделения заряда. Наиболее вероятное объяснение этого явления заключается в том, что небольшой процент центров реакции PSII был фотоингибирован в течение 48-часового процесса восстановления из состояния длительного тушения, что согласуется с измерениями NPQ.
Поразительной особенностью является наличие очень сильного вклада прямой передачи энергии от PSII к PSI в процесс тушения. Другими словами, моделирование показывает, что прямая передача энергии от комплексов PSII к комплексам PSI обеспечивает доминирующий механизм тушения PSII в иглах ранней весны, вызывая выраженное длительное тушение.
Анализ кинетической информации, показанной на 3c, позволяет получить количественную оценку степени эффективности использования света для фотосинтеза и для фотозащиты/тушения, присутствующих как в PSII, так и в PSI.
В летних иглах 70% общей поглощенной энергии приводит к разделению заряда PSII и только 30% — PSI. В SQ иглах процент энергии, поступающей в центр реакции (RC) PSII, падает до 7.1%, в то время как для PSI он увеличивается до 67%. При сильном освещении большинство центров реакции PSII будет закрыто, и часть этой общей энергии, протекающей через RC, может производить вредные реактивные частицы, такие как ROS (или АФК, т.е. активные формы кислорода).
Поскольку процентная доля потока энергии в PSII уменьшается в 10 раз (70% в летних образцах против 7.1% в SQ образцах), это означает, что потенциал окислительного повреждения также снижается в 10 раз в процессе тушения. Это значение увеличивается еще примерно в 2 раза из-за большого количества функционально отделенного и тушенного LHCII. Таким образом, в SQ иглах общее тушение обеспечивает коэффициент защиты 20 для PSII по сравнению с летними иглами.
В образцах ранней весны, прошедших длительное тушение, этот показатель еще выше: лишь 1.5% от общей поглощенной энергии в системе проходит через центр реакции PSII, потенциально вызывая окислительное повреждение (при условии, что все остальные факторы равны). Таким образом, в целом длительное тушение образцов обеспечивает коэффициент защиты примерно 40 для PSII. Столь высокий показатель объясняет способность хвойных растений пережить экстремальные условия длительной зимы и ранней весны.
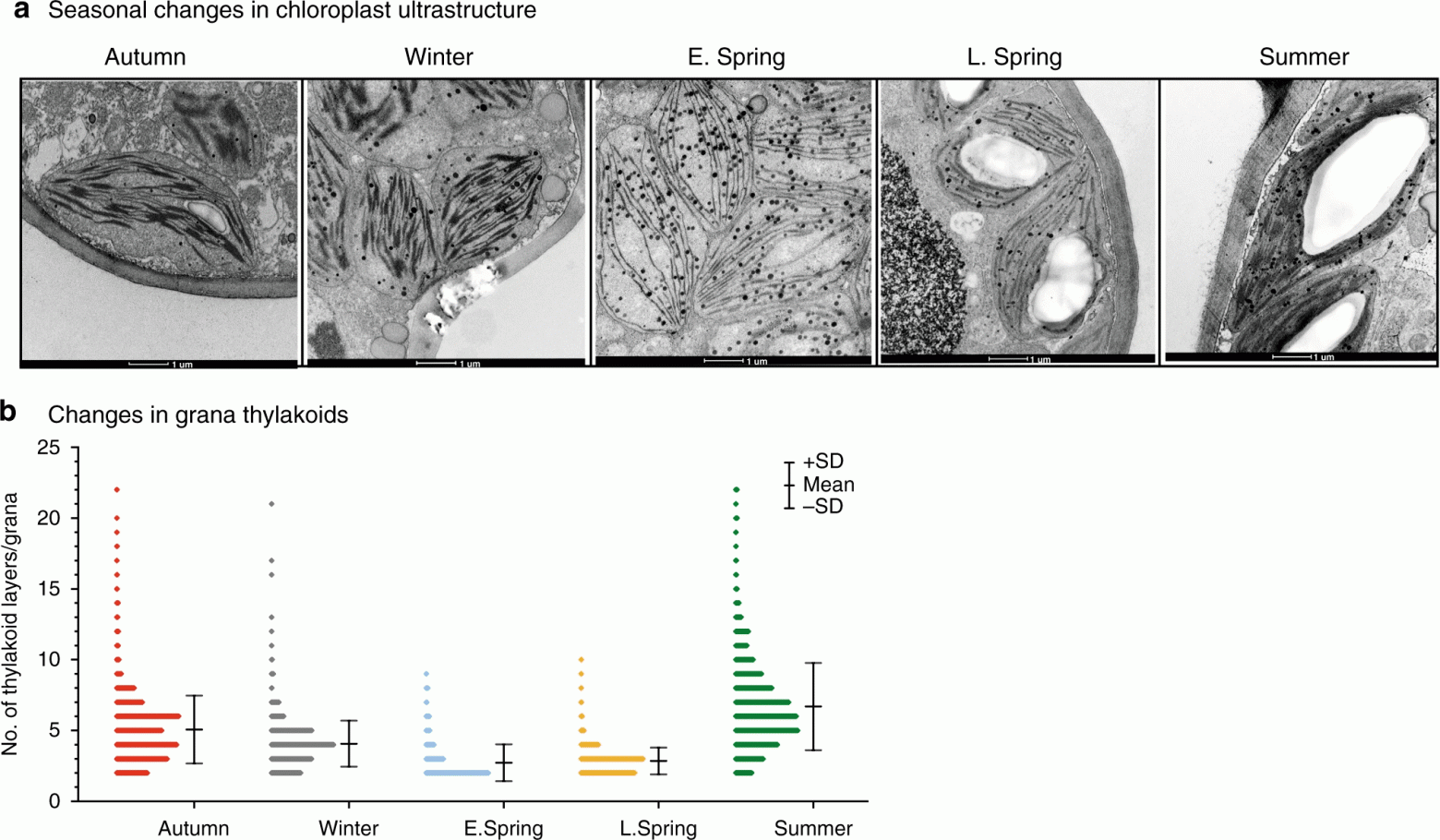
В ходе исследования также наблюдались значимые изменения в структуре гран* хлоропластов (4а).
Изображение №4
Граны* — основные элементы внутренней структуры хлоропласта, представляющие собой стопки из тилакоидов.Структурные изменения показывают, что среднее количество гран на хлоропласт неуклонно снижалось от осени к зиме и достигало минимальных значений ранней весной, т.е. около 68% от летних значений. Удивительно и то, что количество слоев тилакоидов резко снизилось с осени (4.97) до ранней весны (2.72) и снова выросло летом (6.50). Эти изменения подтверждают сдвиг в укладке гран (4b).
Летом и осенью 3-6-слойные стопки гран составляли порядка 60-70: от общего количества стопок. А вот в образцах ранней весны 70-75% составляли двухслойные и 15-20% трехслойные стопки гран (4b). Эти изменения в образцах ранней весны сопровождались кратковременным удвоением количества липидных глобул (пластоглобул) на клетку, когда структура хлоропластов наиболее сильно отклонялась от летнего состояния.
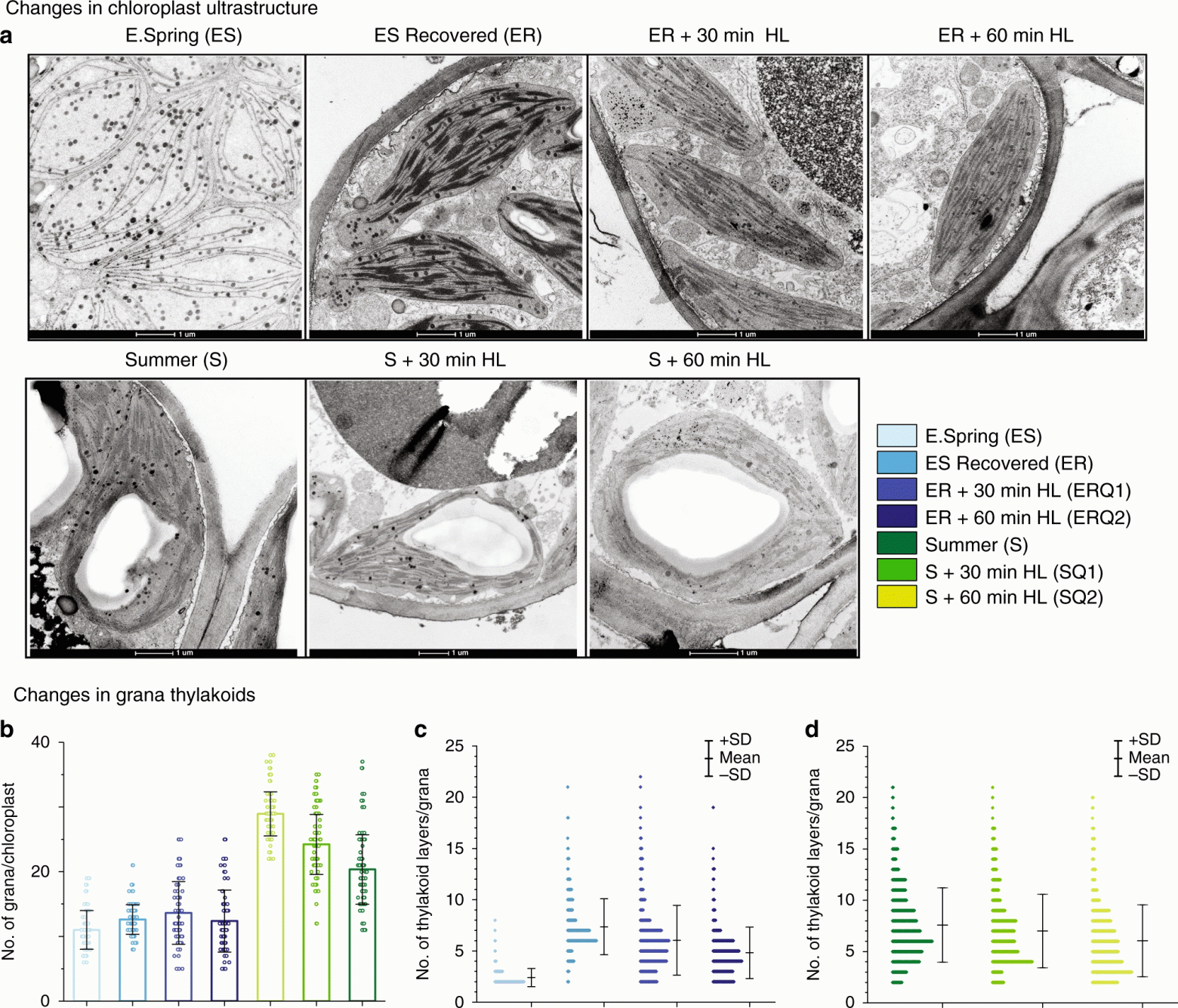
Изображение №5
Далее была проведена электронная микроскопия образцов (собранных летом и ранней весной) после их длительного искусственного восстановления при разных уровнях освещения (5a).
Когда образцы подвергались воздействию света в течение 30 и 60 минут, наблюдалась аналогичная разупаковка гран, как и для образцов ранней весны (5b). Эти изменения в структурах граны, зарегистрированные в образцах (5c и 5d) после кратковременного воздействия освещения, точно подтверждают высокую динамическую пластичность тилакоидной мембраны, которая необходима для адаптации к суровым северным зимам.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
Эволюция это, прежде всего, адаптация к условиям окружающей среды. За долгие тысячи лет адаптации многие растения выработали специфические для вида и для среды обитания механизмы, позволяющие им получать максимальную выгоду с минимальными рисками.
Северные бореальные леса являются весьма экстремальным местом для большинства произрастающих там растений. Самым очевидным фактором «экстремальности», конечно же, является температура. Сильные морозы оказывают особое влияние на процесс фотосинтеза. Зимой световая энергия все же поглощается молекулами хлорофилла, но не может полноценно использоваться в последующих реакциях именно из-за низкой температуры. С наступлением весны ситуация только ухудшается, ибо температура среды все еще низка, а вот солнечного света становится хоть отбавляй. Будучи фактически неактивной система фотосинтеза растения страдает от избыточного излучения, которое может привести к повреждению белковых структур.
Растения обладают двойной системой фотосинтеза, обе части которой работают параллельно и не пересекаются, за счет чего обеспечивается максимальная эффективность фотосинтеза. Зимой структура тилакоидной мембраны. Где и расположены обе системы, меняется. Это приводит к физическому контакту двух фотосистем (PSI и PSII).
В ходе рассмотренного нами сегодня исследования было установлено, что PSII передает часть энергии в PSI, что позволяет защитить хлорофилл в условиях пониженной температуры и/или повышенного солнечного излучения.
По мнению ученых, данный защитный механизм может присутствовать не только в соснах, которые были объектом исследования, но и в других хвойных растениях. Также они уверены, что именно этот адаптационный механизм хвойных растений повлиял на наш вид. Если бы хвойные растения не могли противостоять сильным морозам, то обширные территории в северном полушарии возможно оставались бы долгое время незаселенными. Другими словами, не было бы деревьев — не было бы и древесины, необходимой для постройки жилищ, приготовления пищи, изготовления орудий труда и т.д.
Праздничное послесловие
2020 год подходит к концу. И хоть смена циферки на календаре не имеет особого значения, многие из нас испытывают вполне понятное облегчение. Год уходящий был, мягко (пристойно) говоря, непростой. Хотелось бы сказать, что год приходящий будет в сто крат легче, и что все наладится, воцарится мир во всем мире, а колбаса будет расти на деревьях. Но этого, естественно, не будет. Наш вид не может без препятствий, которые частенько сам себе и создает, а потом героически преодолевает. Возможно наша сила как раз таки в нашей неоднозначности. Мы можем с одинаковой мощью демонстрировать свое невежество и свою гениальность, свой гнев и свою доброту, свое тщеславие и свою скромность. Каждый из нас это совокупность самых разных, самых полярных качеств в нерегулируемом и постоянно колеблющемся процентном соотношении, отними одно — и мы перестанем быть самими собой.
Посему, заканчивая свое разглагольствование, я хочу пожелать всем вам препятствий, которые вы сможете преодолеть, трудностей, которые сделают вас сильнее, и проблем, которые откроют в вас сокрытые таланты. Но, прежде всего, помните, что в этом хаотичном мире, который из года в год ставит новые рекорды безумия, всегда есть кто-то, кому вы дороги.
Благодарю за внимание, оставайтесь любопытствующими, любите друг друга и с наступающим Новым годом, ребята :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Maincubes Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?
