
▎Привет, Хабр!
В этой статье я хотел бы рассказать о том, как с помощью разных современных технологий и фишечек можно решить одну из злободневных проблем современной экологической науки – подсчет численности видов.
| Сразу предупреждаю, я не айтишник. Я зоолог-эколог. Поэтому, в отличие от других подобных статей про ИИ, нейросети и распознавание образов, в этой взгляд будет с точки зрения эколога и того, как мы понимаем данную проблему. И поскольку я изучаю бурых медведей, то речь, главным образом, пойдет про них. |
Современному человеку этот вопрос может вообще показаться странным. Зачем, собственно, считать животных в принципе? Но любой эколог вам объяснит, что численность – одна из самых главных характеристик популяции. Без нее практически невозможно предпринимать какие-либо меры по ее охране. Если речь идет о редких видах, состояние которых находится под угрозой, то численность вообще крайне необходима, потому что на счету буквально каждая особь.
Как раз поэтому многие современные методы оценки численности применяются в первую очередь для редких видов животных. Это же касается и фотоловушек. Думаю, никому не нужно объяснять, что фотоловушки – это такие автономные камеры, которые реагируют на «тепло в движении» и производят фото- или видеосъемку.
| Еще в середине 90-х годов прошлого века были придуманы первые методики учета редких млекопитающих при помощи этих приборов. Стать первыми повезло бенгальским тиграм из Индии. Да, они редкие, а еще очень красивые и, разумеется, котики, и все с этим связанное. Но главным было то, что у тигров есть полоски. И полоски эти настолько уникальны в своей форме и положении, что их узор является, по сути, паспортом каждой особи. |
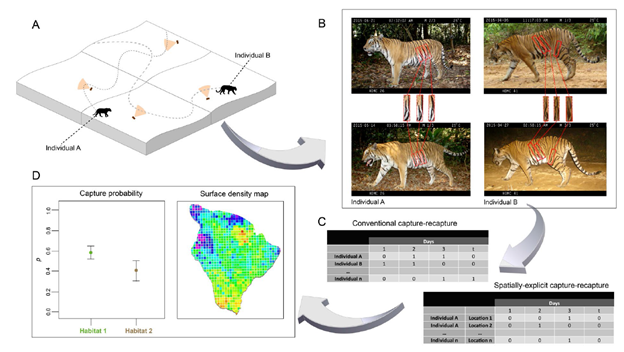
Рис. 1. Упрощенная схема работы методов «отлова-повторного отлова» на примере тигров. Взято из Wearn & Glover-Kapfer (2017).
С редкими кошками все вроде понятно. Но что делать с нередкими и не кошками? С лосями, лисицами, зайцами, ламами, ежиками, куницами и медведями? Ведь других-то видов млекопитающих куда больше, чем кошачьих. И их тоже надо как-то считать. И как назло, ни у кого из них нет ни пятен, ни полосок…
Вот тут-то наша прогрессивная наука и встала в ступор. Но на то она и прогрессивная, что выход все же нашла благодаря прогрессу. Вернее, прогресс пришел немного позже. Сначала пришлось позаимствовать кое-что из других областей знаний. Такая уж она, эта экология – любит понатаскать в себя всего и отовсюду, а потом строить на этом свои теории. И как часто это бывает – решение пришло из физики. А если быть точнее – термодинамики.
| Еще из школьного курса мы помним (хотя бы в общих чертах), что есть такая молекулярно-кинетическая теория идеального газа. В двух словах напомню вам ее постулаты. |
- Размеры частиц (молекул) крайне малы по сравнению с расстояниями между ними;
- Силы притяжения между молекулами не учитываются, а импульс передается только при соударениях;
- Соударения частиц между собой и стенками абсолютно упруги;
- Количество молекул в газе велико и фиксировано;
- Газ находится в состоянии термодинамического равновесия (Ландау и др., 1965).
▎Казалось бы, как эта теория может помочь нам посчитать медведей или куниц?
Тут-то экологи и подсуетились. Один прекрасный британский ученый (в самом деле) по имени Маркус Роуклифф позаимствовал эти постулаты и применил их для фотоловушек и зверушек. Справедливости ради, стоит сказать, что экологи уже давно взяли теорию идеального газа на вооружение. Роуклифф ее просто адаптировал под свои нужды.
На основе этой теории Маркус и его коллеги разработали «модель случайных столкновений» (Random Encounter Model, REM), которая определяет характер контактов («столкновений») между животными и фотоловушками, а после в 2008 г. опубликовали на радостях про это статью (Rowcliffe et al., 2008).
| Действительно, участок пространства с расставленными фотоловушками можно представить как двумерную модель идеального газа, где одни подвижные частицы (особи животных) «случайным» образом двигаются относительно других неподвижных частиц (фотоловушек). При этом «столкновения» (регистрации животных камерами) происходят лишь в эффективной зоне срабатывания фотоловушки (рис.2). |
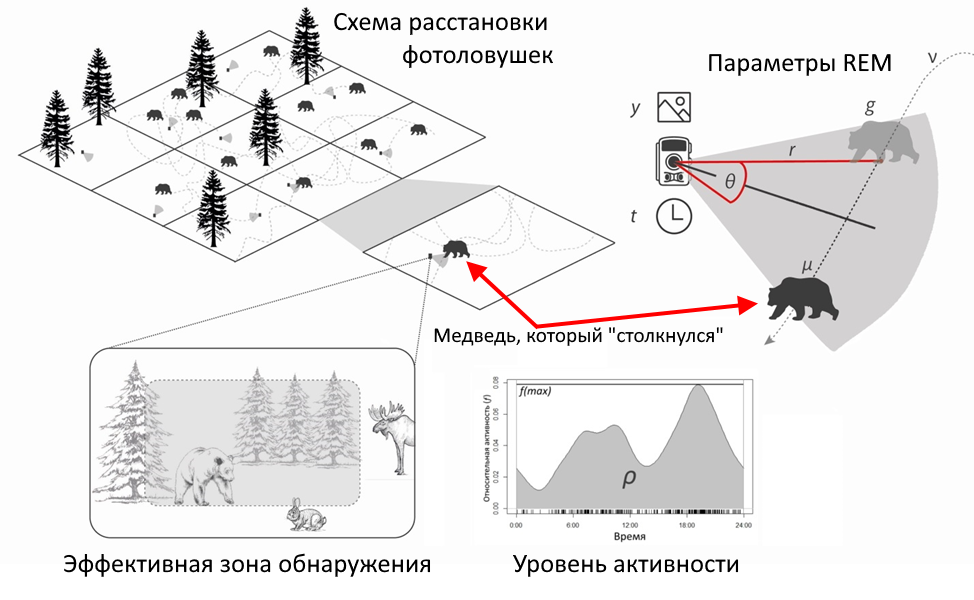
Рис. 2. Принцип работы модели случайных столкновений для расчета численности животных по данным с фотоловушек.
Вероятность «столкновения» животного с фотоловушкой, согласно теории идеального газа, определяется числом самих животных (неизвестная требуемая величина), расстоянием, которое проходят животные за определенное время (чем больше расстояние, тем больше вероятность «столкновения»), и шириной эффективной зоны срабатывания фотоловушки (чем шире зона, тем больше вероятность «столкновения»).
Расстояние, преодолеваемое животным (например, его суточный ход, т.е. расстояние, которое животное проходит за сутки), является производным скорости передвижения и времени, которое оно находилось в активном состоянии (т.е. когда не спало и не отдыхало). Эффективная зона срабатывания фотоловушки представляет собой сектор, определяемый двумя характеристиками – эффективным радиусом (т.е. расстоянием от пассивного инфракрасного (Passive Infra-Red, PIR) датчика камеры до животного) и эффективным углом (т.е. двойным углом между линией до животного (радиусом) и перпендикулярной линией от PIR-датчика) (рис. 3).

Рис. 3. Принцип работы пассивного инфракрасного (PIR) датчика на фотоловушке
Датчик улавливает ИК-излучение в ряде секторов, которые пересекают животные (А.) В основе срабатывания лежит замыкание пироэлемента путем возмущения разницы в ИК-излучении («температуры» в области сектора чувствительности (t) и вне ее (B). Два самых крайних сектора по бокам (1 и 3) и определяют эффективную зону срабатывания фотоловушки (С). Взято из Wearn & Glover-Kapfer (2017).
Все математические подробности самих вычислений мы сейчас опустим. Сейчас нас интересует два главных параметра, которые входят в REM – эффективная зона срабатывания фотоловушки и длина суточного хода зверя, которого мы хотим посчитать. Вернее сказать, эти параметры интересуют не только нас, а вообще очень многих (уж поверьте на слово).
Именно расчет эффективной зоны фотоловушки и длины суточного хода долгие годы представлял сложности для ученых по всему миру. Вот как их рассчитывать? Паспортные характеристики самой камеры (дальность срабатывания и угол обзора объектива) не совпадают с реальным радиусом и углом PIR-датчика. Для определения этой зоны ученые изощрялись, как могли.
Тащили в леса ноутбуки и перед фотоловушкой уже по фотографиям на экране лэптопа смотрели, где был зверь, потом прикидывали это место и с помощью компаса и линейки измеряли угол и радиус. Другие привязывали ленточки на деревья или примечали какие-то объекты (камни, кустики и т.п.), до которых заранее измеряли расстояния (рис. 4). Так худо-бедно, но эффективную зону удавалось рассчитать.

Рис. 4. Пример измерения эффективного радиуса ® и угла срабатывания фотоловушки (2ɑ) в исследовании численности различных копытных в Швеции с помощью красных ленточек. Взято из Pfeffer et al. (2018).
▎А как же быть с суточным ходом?
По фотографиям-то его уж точно никак не рассчитаешь! Остаются либо чьи-то экспертные данные из других статей, либо GPS-телеметрия (которая очень сильно этот самый ход занижает и вообще жутко дорогая). Короче, проблем было много. Кто-то, попробовав и поползав на брюхе пару месяцев в дождевом лесу Эквадора, плюнул на этот ваш REM и пошел дальше считать своих котиков. Кто-то набрался терпения и стал ждать, пока самые умные что-то придумают. И вот как раз-таки самые умные взяли и придумали. А пока что для всех была очень удобная и в тоже время вполне оправданная отмазка Говарда Старка (рис. 5).

Рис. 5. Говард Старк как бы намекает, что он пытался, но не фортануло.
И вот как раз тут подключился уже упомянутый выше «прогресс». Хоть и с заметным опозданием. Но в любом случае, где-то в 2020 г. был разработан простенький алгоритм компьютерного зрения, который враз решил все проблемы. И разработал его наш старина Маркус со своими друзьями. Вокруг этого алгоритма буквально на коленке собрали минималистичное веб-приложение (называется Animal-Tracker), и наступило повальное счастье (ну, у меня так точно).
| С помощью Animal-Tracker удается переводить разницы координат пикселей на фотографиях во вполне реальные метрические длины. Насколько реальные – зависит уже от качества калибровки. А калибровка в данном случае очень простая (хоть и муторная). Для этого нужна любая ровная палка (по научному – калибровочный шест), размеченная на отрезки по 20 см. |
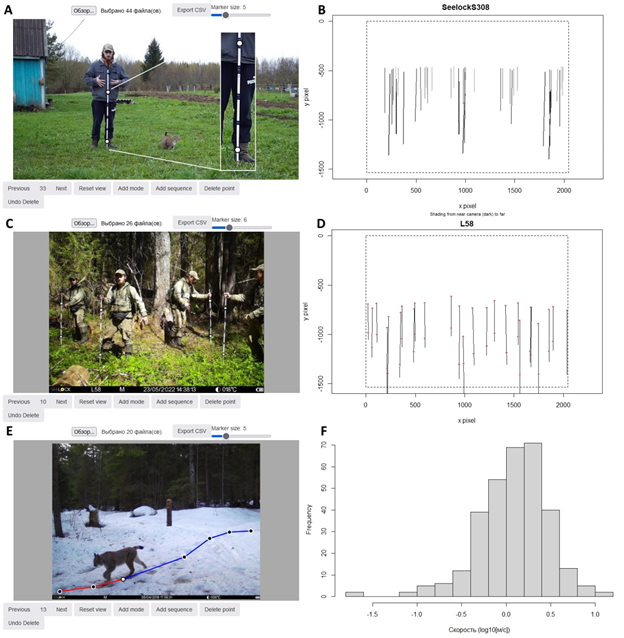
Рис. 6. Калибровка фотоловушек
Пример калибровки модели фотоловушки (A, B), места установки фотоловушки (С, D), воссоздания пройденного пути животного на изображениях (Е) при помощи веб-приложения Animal-Tracker, а также гистограмма распределения десятичных логарифмов рассчитанных скоростей (F). На фото A я тыкаю палку перед фотоловушкой и пальцами показываю, сколько до нее метров. Рядом мой кот следит, чтобы я делал все правильно. Потом уже на этапе обучения модели я отмечаю крайние насечки на самой фотографии точками. В результате модель воссоздает мои палки в виде вполне реальных геометрических объектов (B, D). На рисунке E показан уже не мой кот, для которого таким образом можно рассчитать пройденное расстояние в кадре.
Но это еще не все. Добрый дядя Маркус пошел дальше и решил, что с этой же помощью можно рассчитать и суточный ход. Ведь если мы теперь можем измерять линейные размеры на фотографиях, значит, мы можем посчитать, сколько медведь прошел в кадре. Для этого на каждой фотке с медведем помечаем точкой проекцию его центра масс на земле. Потом соединяем все точки в общей серии фоток – вот мы и получили ломаную линию, которая ни что иное как восстановленный путь медведя (рис. 7 и еще 6E)! Время каждой фотки достаем из ее метаданных, а значит, знаем за сколько медведь этот путь прошагал. В результате получаем скорость передвижения. Ну, а дальше простым умножением на уровень активности (долю времени суток, когда медведь находится в движении) получаем и сам суточный ход.
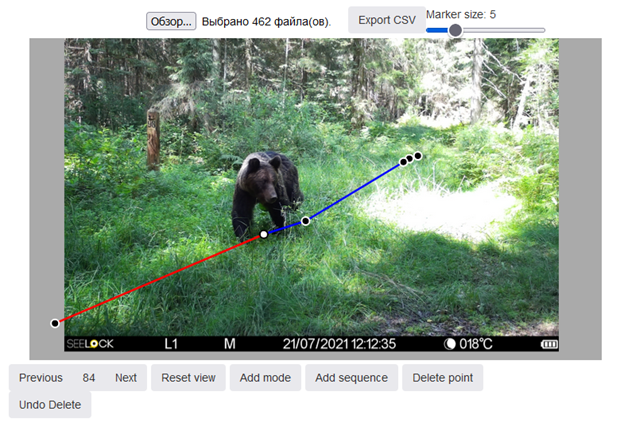
Рис. 7. Пример воссоздания пройденного пути бурым медведем по кадрам фотоловушки в интерфейсе веб-приложения Animal-Tracker. Синие сегменты – путь пройденный, красные – путь предстоящий. Точки – проекции центра массы зверя на поверхность земли. Взято из Огурцов (2023).
▎Тем временем не один дядя Маркус что-то придумывал.
Горячие испанские ребята из команды Пабло Паленсиа подумали и решили, что просто так рассчитывать суточный ход – это не комильфо и вообще как-то слишком просто. Надо все как-то усложнить, а заодно и объяснить по-научному. И поняли они, что скорость медведей-то непостоянна и зависит от их поведения. Вот он травку жует – скорость маленькая, а тут за лосем гонится – скорость большая. И чаще-то медведи спокойно ходят, нежели носятся как ошпаренные по 25-30 км/ч.
Поэтому среднюю арифметическую считать нехорошо, выборка-то сильно скошенная в сторону небольших скоростей получается. Значит, скорость нужно рассчитывать сообразно с поведенческим паттерном (рис. 8). В итоге Пабло и друзья подсуетились и запилили модель машинного обучения, которая сама находит разные паттерны (методом кластеризации k-средних), считает для них свои скорости и уровни активности, а потом выводит итоговый суточный ход. Модель эту назвали страшными зубодробительными буквами DRRB (Day Range estimation with Ratios between Behaviors).
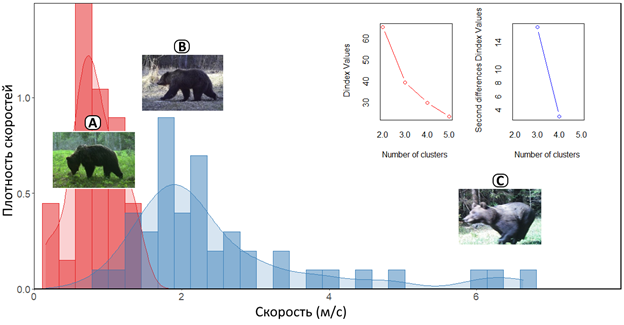
Рис. 8. Пример расчета показателя скорости передвижения медведя для выделенных поведенческих паттернов на основе моделирования с помощью DRRB.
В данном случае модель выделила 2 кластера (A – медленный, соответствующий, в основном, питанию и исследованию; B – средний, соответствующий перемещениям между местообитаниями). Также заметны своеобразные «выбросы» (наиболее высокие скорости, соответствующие погоням медведей за лосями, но их число недостаточно для выделения третьего отдельного кластера C. Взято из Огурцов (2023).
В итоге прогрессивные британо-испанские умы решили почти все проблемы, да еще и вдобавок понаписали кучу всяких R-скриптов и выложили все на гитхаб. Пользуйтесь на здоровье и считайте своих медведей, белочек и прочих ежиков. И все бы хорошо, но… тут снова вмешались физики. Они-то ребята хоть и веселые, но при этом во всем опираются на скучную теорию. Это экологи любят быстрее в лес смотаться да кое-как данные пособирать.
А физики с математиками на пару вечно им в этом мешают. REM основана на теории идеального газа? Да. Значит, должны соблюдаться ее постулаты, если ты и вправду хочешь посчитать своих медведей. И если большинству постулатов с грехом пополам удается как-то следовать, то вот одному из них – ну прям вообще никак. И кроется он в самом названии метода – модель СЛУЧАЙНЫХ столкновений.
Означает это то, что наши медведи должны двигаться СЛУЧАЙНО относительно фотоловушек. А поскольку медведи случайно никогда и нигде не двигаются, то значит, сами фотоловушки должны быть случайно расставлены по территории. Ну, грубо говоря, создать в ГИС набор рандомных точек, а потом прийти по GPS на каждую и повесить там фотоловушку. Глухой лесной бурелом? Что поделать… Топкое низинное болото? Иначе нельзя… Отвесный утес, где вчера сорвались двое скалолазов? Значит, наука (а точнее физики) требует новых жертв… Ну, на самом деле все не так радикально и каждую случайную точку можно корректировать на 100-200 м, но суть вы уловили. Фотоловушки реально должны стоять рандомно, т.е. как раз «просто так».

Первый меч Гондора пытается разобраться в принципе случайных столкновений.
Проблема вся в том, что рандомно фотоловушки никто (почти никто) не ставит. Потому что это овер много денег. Ведь медведи-то по топким болотам и лесным буреломам очень редко ходят. Они больше по дорожкам да тропинкам. Поэтому, чтобы их «поймать» на фотоловушки в более-менее приличном для статистики количестве, нужны сотни камер. А столько есть только у самых избранных и богатых научных коллективов. И вот тогда на помощь пришел наш третий супергерой – Херман Гаротте (опять испанец! Хм…).
| Он придумал элементарный способ избежать случайного дизайна исследований – специальный поправочный коэффициент, который корректирует нашу REM и позволяет работать с фотоловушками на тропах и дорогах. Браво, Херман! |
▎Какой у нас сухой остаток?
В следующем году у REMа юбилей – 15 лет. И только за последние 3 года он реально развился до того уровня, что стал доступен всем экологам, кто более-менее владеет программированием в R и понимает, куда нужно тыкать своими палками (прошу прощения, калибровочными шестами). Теперь у нас есть вся теория Роуклиффа-Паленсио-Гарроте в виде научных статей в крутых международных журналах, практика в виде кодов на гитхабе и никаких преград для исполнения мечты почти всех фотоловушников мира – посчитать своих любимых зверушек по фоточкам.
| Мы сейчас не будем говорить о пресловутых нейросетях, которые нам помогают этих зверушек на этих самых фоточках искать – это уже и так все знают и умеют. Также ничего не скажем про те же сети, которые считают моржей/тюленей с дронов. Тут нет ничего сложного. Другое дело – посчитать тех зверей, кто живет в лесу, и вряд ли ты их с дрона когда увидишь. Для этого нужны и тяжелые полевые работы, и нейросети, и компьютерное зрение, и машинное обучение. Поэтому реально посчитать медведей у себя в заповеднике мне удалось только в этом году. Для меня это был прям новый этап в wildlife biology какой-то. Так долго все это казалось недоступным. |
Использованная литература
Ландау Л.Д., Ахиезер А.И., Лифшиц Е.М. 1965. Курс общей физики: Механика. Молекулярная физика. М.: Наука. 405 с.
Огурцов С.С. 2023a. Оценка плотности населения млекопитающих с помощью фотоловушек на основе модели случайных столкновений: теоретические основы и практические рекомендации // Nature Conservation Research. Заповедная наука. Т. 8(1). С. 1-23. DOI: 10.24189/ncr.2023.007
Огурцов С.С. 2023b. Оценка плотности и численности популяционной группировки бурого медведя с помощью фотоловушек в Центрально-Лесном заповеднике (запад Европейской России) // Nature Conservation Research. Заповедная наука. Т. 8(2). DOI: 10.24189/ncr.2023.008
Garrote G., Pérez de Ayala R., Álvarez A., Martín J., Ruiz M., De Lillo S., Simón M. 2021. Improving the random encounter model method to estimate carnivore densities using data generated by conventional camera-trap design // Oryx. Vol. 55(1). P. 99–104. DOI: 10.1017/S0030605318001618
Palencia P., Fernández-López J., Vicente J., Acevedo P., 2021b. Innovations in movement and behavioural ecology from camera traps: day range as model parameter. Methods of Ecology and Evolution. Vol. 12(7). P. 1201–1212. DOI: 10.1111/2041-210X.13609
Pfeffer S.E., Spitzer R., Allen A.M., Hofmeester T.R., Ericsson G., Widemo F., Singh N.J., Cromsigt J.P.G.M. 2018. Pictures or pellets? Comparing camera trapping and dung counts as methods for estimating population densities of ungulates // Remote Sensing in Ecology Conservation. Vol. 4. P. 173–183. DOI: 10.1002/rse2.67
Rowcliffe J.M., Field J., Turvey S.T., Carbone C. 2008. Estimating animal density using camera traps without the need for individual recognition // Journal of Applied Ecology. Vol. 45. P. 1228–1236. DOI: 10.1111/j.1365-2664.2008.01473.x
Wearn O.R., Glover-Kapfer P. 2017. Camera-trapping for conservation: a guide to best-practices. WWF Conservation Technology Series 1(1). WWF-UK, Woking, United Kingdom. 181 p.
Огурцов С.С. 2023a. Оценка плотности населения млекопитающих с помощью фотоловушек на основе модели случайных столкновений: теоретические основы и практические рекомендации // Nature Conservation Research. Заповедная наука. Т. 8(1). С. 1-23. DOI: 10.24189/ncr.2023.007
Огурцов С.С. 2023b. Оценка плотности и численности популяционной группировки бурого медведя с помощью фотоловушек в Центрально-Лесном заповеднике (запад Европейской России) // Nature Conservation Research. Заповедная наука. Т. 8(2). DOI: 10.24189/ncr.2023.008
Garrote G., Pérez de Ayala R., Álvarez A., Martín J., Ruiz M., De Lillo S., Simón M. 2021. Improving the random encounter model method to estimate carnivore densities using data generated by conventional camera-trap design // Oryx. Vol. 55(1). P. 99–104. DOI: 10.1017/S0030605318001618
Palencia P., Fernández-López J., Vicente J., Acevedo P., 2021b. Innovations in movement and behavioural ecology from camera traps: day range as model parameter. Methods of Ecology and Evolution. Vol. 12(7). P. 1201–1212. DOI: 10.1111/2041-210X.13609
Pfeffer S.E., Spitzer R., Allen A.M., Hofmeester T.R., Ericsson G., Widemo F., Singh N.J., Cromsigt J.P.G.M. 2018. Pictures or pellets? Comparing camera trapping and dung counts as methods for estimating population densities of ungulates // Remote Sensing in Ecology Conservation. Vol. 4. P. 173–183. DOI: 10.1002/rse2.67
Rowcliffe J.M., Field J., Turvey S.T., Carbone C. 2008. Estimating animal density using camera traps without the need for individual recognition // Journal of Applied Ecology. Vol. 45. P. 1228–1236. DOI: 10.1111/j.1365-2664.2008.01473.x
Wearn O.R., Glover-Kapfer P. 2017. Camera-trapping for conservation: a guide to best-practices. WWF Conservation Technology Series 1(1). WWF-UK, Woking, United Kingdom. 181 p.
Автор: Сергей Огурцов, зоолог
НЛО прилетело и оставило здесь промокод для читателей нашего блога: — 15% на все тарифы VDS (кроме тарифа Прогрев) — HABRFIRSTVDS

