
Мы подошли к моменту, когда от пересказа азов нейробиологии и теории нейронных сетей нам предстоит перейти к тому новому, что содержит предлагаемая модель. Тем, кто только приступил к чтению цикла я советую начать с первой части.
Вернемся к описанию работы реальных нейронов. Сигналы от одних нейронов через их аксоны поступают на входы других нейронов. В химических синапсах происходит выброс медиатора, который в зависимости от типа синапса оказывает либо активирующее, либо тормозящее воздействие на принимающий сигнал нейрон. Чувствительностью синапса, которая может меняться, определяется вклад этого синапса в общее возбуждение. Если суммарное воздействие превышает определенный порог, то происходит деполяризация мембраны и нейрон генерирует спайк. Спайк – это одиночный импульс, продолжительность и амплитуда которого не зависит от того, какая синаптическая активность его породила.
Самая простая модель, навеянная импульсной активностью нейрона, – это модель порогового сумматора. При этом, исходя из того, что спайк можно сопоставить с бинарным сигналом, полагают, что входы и выход сумматора принимают значения только 0 и 1. Если на входы такого формального нейрона подавать импульсную картину, повторяющуюся от такта к такту, то нейрон в зависимости от настройки его весов должен либо каждый такт выдавать ответный сигнал, либо молчать. Это вполне логично – постоянная входная картина соответствует постоянному результату на выходе.
Если попытаться приблизить модель порогового сумматора к реальности, то первое, что придется сделать – это допустить, что картина входной активности может быть не строго синхронна. То есть сигналы на разных входах могут кодироваться импульсами, имеющими каждый свою частоту. При таком допущении уже нельзя просто использовать мгновенную картину состояния входов. Потребуется подобрать показательный временной интервал и пользоваться накопленной за этот интервал картиной активности. При разных частотах входных сигналов на некоторых интервалах входные импульсы будут образовывать кучности, достаточные для активации нейрона, а на других давать разряжения, оставляя нейрон неактивным. Таким образом, ответ нейрона приобретет собственную частоту, которая будет зависеть от частоты входных сигналов и чувствительности соответствующих синапсов.
Такие рассуждения приводят нас к модели нейрона как линейного сумматора, в которой уровень сигналов на входах нейрона и его ответ описываются не двумя уровнями, а скалярными величинами, которые соответствуют частотам следования спайков. Переход к линейному сумматору позволяет сильно упростить моделирование и отчасти забыть про изначальную химическую природу нейронной активности.
Но все имеет свое начало. Чтобы информация поступила в мозг, необходимы нейроны, взаимодействующие с внешним миром. Чувствительность такого взаимодействия резко отличается от синаптической чувствительности. В синапсах, количество которых у одного нейрона может измеряться десятками тысяч, медиатор высвобождается из пузырьков, имеющих постоянную емкость. Минимальный квант выброса медиатора – это порция из порядка 7 000 молекул. Сенсорные нейроны работают совсем с другими объемами. Так, зрительные палочки активируются буквально от двух квантов света, нейроны-рецепторы обонятельной системы способны обнаружить всего несколько молекул пахучего вещества. Столь высокая восприимчивость достигается за счет механизмов внутреннего усиления сигнала.
Взаимодействие нейрона с окружающей средой происходит за счет белковых молекул – рецепторов, которые позволяют внешнему химическому воздействию изменять состояние внутри клетки. Вещество, которое взаимодействует с определенным типом рецепторов, называют их лигандом. Для синаптических рецепторов лиганды – это те самые нейромедиаторы, что обеспечивают взаимодействие нейронов.
Соседние рецепторы могут соединяться, создавая димеры (рисунок ниже), которые в свою очередь, объединяясь, формируют рецептивные кластеры.

Кластеризация рецепторов. А – одиночный рецептор и его взаимодействие с окружающими рецепторами. Б – мономерная рецептивная молекула. В – рецептивный димер. Г – объединение двух мономеров в контактный (Д) и комбинационный (Е) димеры. (Радченко, 2007)
Рецепторы делятся на ионотропные и метаботропные. Ионотропные рецепторы создают ионные каналы, которые перемещают заряженные ионы через мембрану, меняя мембранный потенциал. Когда лиганд взаимодействует с ионотропным рецептором, последний меняет проводимость ионного канала, открывая или закрывая его. Именно такие рецепторы расположены в синаптических щелях. Их совместная работа определяет, быть или не быть спайку, вызванному сложением внешних сигналов.
Несколько иная природа у метаботропных рецепторов. Они располагаются за пределами синапсов и не создают ионных каналов. При взаимодействии со своим лигандом они запускают работу внутриклеточных посредников, которые обеспечивают усиление исходного сигнала в сотни тысяч раз, что в итоге приводит к активации нейрона. Это означает, что даже малая доля вещества, которое является лигандом для метаботропного рецептора, может вызвать у соответствующего нейрона спайк. Такой механизм используют сенсорные нейроны, именно он позволяет им реагировать на ничтожные сторонние воздействия. Но кроме сенсорного восприятия метаботропные рецепторы отвечают и за существенную долю мозговой активности вообще.
Характерный стимул для нейрона – это такая картина входных сигналов, которая совпадает с картиной чувствительности его синапсов. Чем ближе входной сигнал к характерному стимулу, тем выше частота спайков, генерируемых нейроном. Эту реакцию принято называть вызванной активностью. Она возникает у всех тех нейронов, которые как-то отреагировали на текущий образ. Но вызванная активность составляет лишь незначительную часть общей активности мозга. Основная же активность приходится на так называемую фоновую активность (рисунок ниже).

Реакция нейрона на стимул и фоновая (спонтанная) активность
Фоновая активность складывается из возникающих время от времени одиночных спайков. Такие спайки называют спонтанными, так как они проявляются независимо от наличия или отсутствия вызванной активности. Если оградить мозг от внешней информации, спонтанная активность на сенсорных зонах не ослабнет, а наоборот, только усилится. Для отдельного нейрона его спонтанная активность выглядит серией случайных спайков. Но для нейронов одной зоны коры эта активность складывается в общий ритм, что суммарно создает значительные электрические колебания. Именно эти колебания можно зафиксировать, приложив электроды к коже головы, что собственно и называется электроэнцефалографией.
Для разных зон коры и разных состояний человека свойственны свои частоты и уровни таких ритмов. Наиболее сильно ритм от отдельной зоны коры прослеживается на участке кожи, находящемся непосредственно над этой зоной. Поэтому для записи ритмов мозга используют набор электродов, равномерно распределенных по поверхности головы. Результат выглядит как набор графиков, каждый из которых записан от своего электрода (рисунок ниже).

Пример электроэнцефалограммы. Резкое увеличение амплитуды соответствует началу эпилептического припадка
Если вызванная активность – это следствие работы ионотропных рецепторов, то спонтанная активность – результат срабатывания рецепторов метаботропных. Когда импульсы регулярной или спонтанной активности, распространяясь по аксону, доходят до нейрона получателя, они вызывают выброс медиатора в синаптическую щель. Этот медиатор определяет вклад синапса в процесс вызванной активности нейрона. Затем происходит его обратный захват, и синапс восстанавливается до своего исходного состояния. Но часть медиатора выбрасывается за пределы синаптической щели и растекается по пространству, образованному телами нейронов и телами окружающих их глиальных клеток. Это явление называется спиловером от английского spillover – переполнение, перелив. Расположенные поблизости друг от друга синапсы образуют так называемые синаптические ловушки. В этих ловушках тормозные и активирующие медиаторы создают интерференционную картину. Это значит, что каждое сочетание импульсов вызывает пучность медиатора в определенном уникальном месте мембраны. А.Н. Радченко показал, что метаботропные рецептивные кластеры, расположенные в местах таких пучностей, способны менять свои свойства и впоследствии реагировать на повторение той же окружающей импульсной картины (Радченко, 2007).
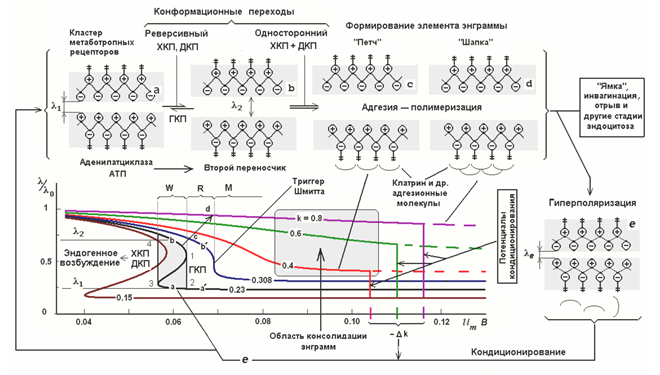
Различные состояния метаботропного рецептивного кластера (Радченко, 2007)
Радченко описал несколько состояний, характерных для метаботропных рецептивных кластеров, и сопоставил эти состояния с различными стадиями запоминания (рисунок выше). Способность рецепторов влиять на состояние клетки зависит от положения их расщепленных концов. Чем больше расстояние между ними, тем сильнее наружные концы выступают на поверхности, а внутренние проникают в среду клетки. Соответственно, большему расстоянию соответствует большая чувствительность рецептора. При уменьшении расстояния концы рецепторов погружаются в мембрану, и рецепторы утрачивают свою способность влиять на состояние нейрона. Расстояние может управляться мембранным потенциалом нейрона. Деполяризация выталкивает концевые части из мембраны наружу и внутрь клетки соответственно, а гиперполяризация встречно втягивает их в мембрану.
Изначально метаботропные рецептивные кластеры обладают гистерезисными характеристиками. Гистерезис подразумевает, что поведение системы определяется ее предысторией (график ниже). Перейдя в определенное устойчивое насыщенное состояние, система не возвращается обратно после снятия усилия, а остается в этом положении. Для выведения ее из этого положения теперь необходимо приложить обратное усилие, которое после перехода определенного порога переведет систему в другое устойчивое состояние, обладающее тем же свойством «залипания».
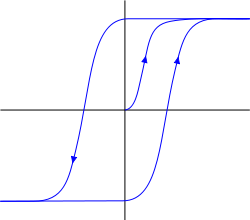
Гистерезис
Гистерезис придает рецепторам свойства триггера. Реагируя на пучность медиатора, рецепторы могут «залипать» как в состоянии вытолкнутости из мембраны, так и в состоянии погружения в нее. Первое состояние говорит о том, что нейрон становится чувствителен к определенной картине окружающей активности и может отреагировать на ее повторение собственным спайком, второе, наоборот, делает его к ней невосприимчивым. Но это залипание носит краткосрочный характер, так как определенное изменение мембранного потенциала может сбросить кластер в исходное состояние.
Радченко сопоставил такое поведение рецептивных кластеров с краткосрочной (кратковременной) памятью. То есть то, что в каждый конкретный момент определяет нашу мысль – это, с одной стороны, картина активности нейронов, а с другой стороны, такая фиксация следов этой активности, которая позволяет быстро восстанавливать недавние картины. Недолговечность этих следов обеспечивает оперативность памяти. Мы можем быстро вернуться к мысли, которая была секунду или минуту назад, но теряем эту легкость, когда проходят часы или сутки.
Под воздействием медиаторов и сильного сдвига мембранного потенциала рецепторы могут терять гистерезисные свойства и переходить в стабильный режим, который надолго фиксирует приобретенное состояние кластера, что можно сопоставить с долговременной памятью. При этом в зависимости от положения фиксации, вытолкнутое состояние или утопленное, кластер либо реагирует на определенную картину активности, либо наоборот – приобретает стабильную нечувствительность к ней. Эти устойчивые состояния могут храниться очень долго, можно сказать вечно, хотя при определенных условиях и они могут быть перезаписаны.
Далее в нашей модели мы будем исходить из того, что нейроны мозга обладают тремя формами хранения информации. Одна форма – это изменение чувствительности синапсов, что определяет характерный для нейрона образ. Вторая форма – это кратковременная триггерная фиксация на метаботропных рецептивных кластерах картины окружающей активности. И третья – это долговременная фиксация, за счет перехода рецептивных кластеров в устойчивое состояние, части кратковременно зафиксированных картин.
Чувствительность синапсов нейрона описывает единственный характерный для него образ (если не учитывать нейроны с несколькими секциями синапсов). При этом внесинаптическая поверхность нейрона, которая содержит сотни тысяч метаботропных рецептивных кластеров, может хранить сопоставимое число воспоминаний, указывающих, в каких событиях проявлялся этот характерный образ.
Обратите внимание, что импульсная активность нейронов складывается из двух составляющих. Одна составляющая – это картина вызванной активности, которая говорит о том, какие нейроны узнали характерные для них стимулы. Вторая составляющая – это картина условно спонтанной активности, которая существует относительно независимо от первой, и которая создает общий фон работы мозга. Этот фон менее акцентирован, чем вызванная активность, но более масштабен, так как распространен по всему пространству коры.
Не побоюсь сказать, что именно фоновая активность — одна из самых больших загадок мозга. В следующей части мы попробуем дать разгадку этой тайны.
Продолжение
Если где-то слишком кратко, непонятно или невнятно изложено, пожалуйста, отпишитесь в комментариях. У меня уже глаз настолько замылен, что мне трудно поставить себя на место того, кто первый раз воспринимает этот материал. При этом, если не трудно, укажите уровень вашего общего знакомства с темой.
Предыдущие части:
Часть 1. Нейрон
Часть 2. Факторы
Часть 3. Персептрон, сверточные сети

